Der letzte Abschnitt behandelte die Kompartimentierung von Hyperzyklen zur Auswahl der tauglichsten Gene, die letztendlich einen genetischen Apparat samt Replikation und Translation konstruieren sollten. Die Abgrenzung von der Umwelt ging mit den beiden Problemen der Umhüllungssubstanz und, verbunden damit, dem (geregelten) Stofftransport durch diese Hülle einher. Doch bevor wir uns mit der Umhüllung direkt beschäftigen, möchte ich einige interessante Experimente und Entdeckungen beschreiben, die Gedankengänge in dieser Richtung maßgeblich beeinflussen können.
Alexander I. Oparin: Koazervathypothese
Der russische Forscher Oparin experimentierte in seinem Leben mit Makromolekülen, die in wässrigen Lösungen kleine kugelförmige Tröpfchen ergaben. Er untersuchte weiterhin, wie die Tröpfchen es schaffen stabil zu bleiben. Seine Theorie war, dass der Stoffwechsel eines Protobionten der Ausgangspunkt des Lebens war und der genetische Apparat später als "Kleinigkeit" dazukam.
Diese Tröpfchen nennt man Koazervate und diese schwimmen dann in dem wasserreichen Medium. Ihre Größe beträgt etwa 1 - 500 µm (Mikrometer). Diese Koazervate sind durch eine Art Membran, einer Verdickung an der Außenwand, von der Außenwelt abgeschirmt. Er verwendete verschiedene Substanzenkombinationen um Koazervate zu erhalten:
- Proteine und Kohlenhydrate: Histon, gummi arabicum,
- zwei Proteine: Histon und Albumin, sowie
- Proteine und Nucleinsäuren: Histon (Clupein) und DNS (RNS).
Hier muss man allerdings anmerken, dass diese Stoffe nicht gerade die primitivsten ihrer Art, sondern vielmehr Produkt des Zellstoffwechsels und der Evolution sind, sodass diese Versuche in dieser Konstellation allenfalls als Modell dienen können. Einige dieser Koazervate sind unbeständig und sinken unter Bildung einer wasserfreien Schicht auf den Boden. Oparin suchte danach nach Möglichkeiten, wie er diese Koazervate für einige Stunden oder Wochen stabil halten könnte. Erstaunlich dabei ist, dass eben dieses durch einen primitiven Stoffwechsel erreicht werden kann.
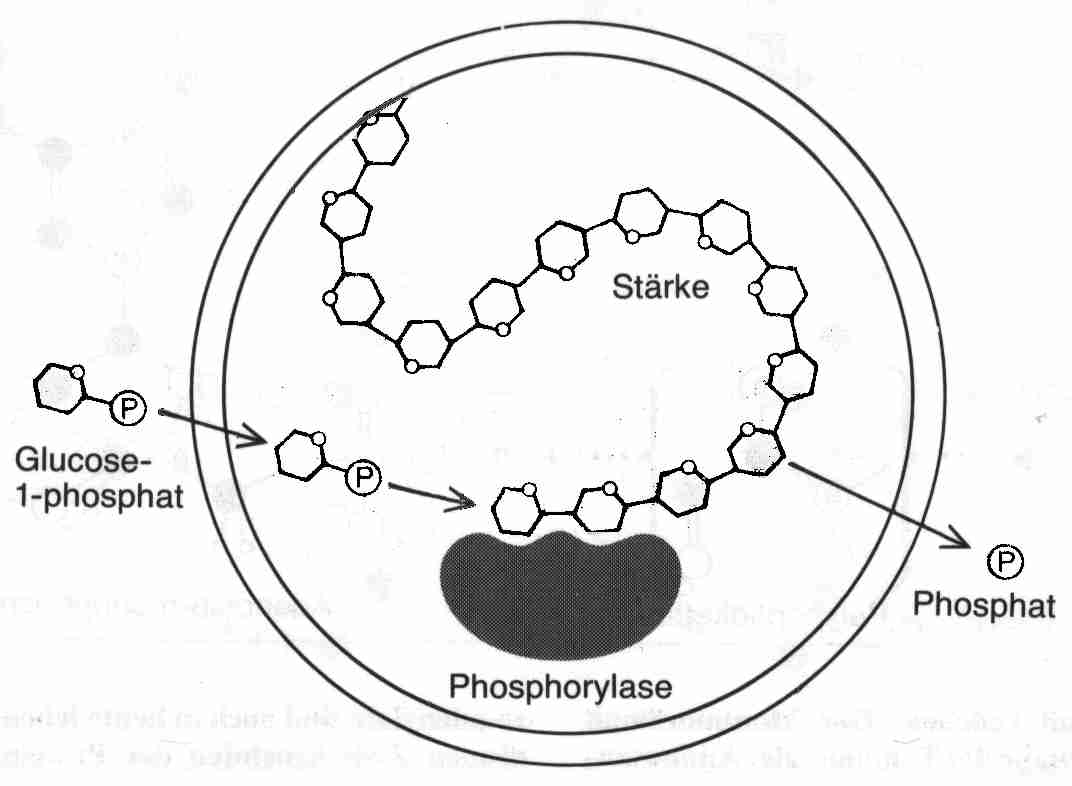
Oparin versetzte seine Lösung mit dem Enzym Phosphorylase, das in die Tröpfchen hineindiffundierte. Als Nahrung bot er diesem Stoffwechsel Glucose-1-phosphat an. Diese wurde unter Abspaltung der Phosphatgruppe unter Nutzung dieser energiereichen Bindung zu Stärke polymerisiert. Das Phosphat wurde als Abfallprodukt abgegeben, die Stärke aber trug zum Wachstum des Koazervats bei. Wenn die Tröpfchen zu groß wurden, zerfielen sie in kleinere Tochtertröpfchen. Wenn diese Tochtertröpfchen das Enzym in sich trugen, wuchsen auch sie, allerdings langsamer. Die Enzymkonzentration in den Tröpfchen wurde immer kleiner, je öfter sie sich teilten und irgendwann hörten sie auf zu wachsen. Hier fand Oparin sicher auch seinen Ansatz, dass der genetische Apparat nur dafür zu sorgen hatte, dass die Tochtertröpfchen das Enzym produzieren konnten. Dieses Koazervat, dass sie auch in der unteren Abbildung 1.1 sehen können, hat einen einstufigen Stoffwechsel.
 |
 |
 |
| Abb. 1.1 | Abb. 1.2 | Abb. 1.3 |
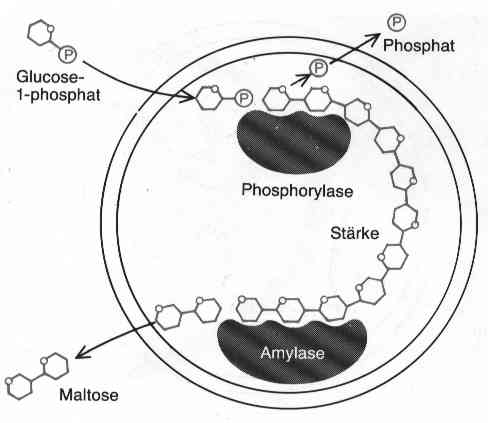
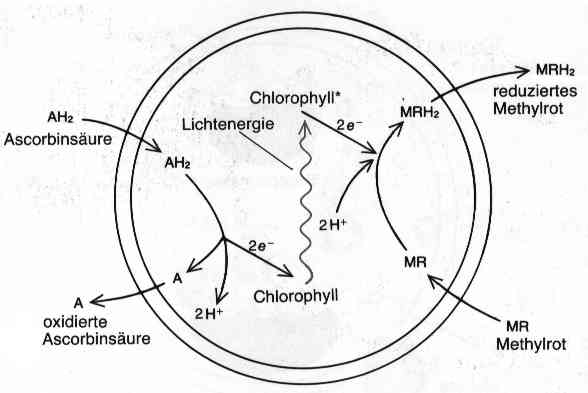
Ähnliche Anordnungen (Abb. 1.2) zeigten, dass auch Koazervate mit mehrstufigem Stoffwechsel wachsen und sich vermehren konnten. Doch Oparin ging noch weiter und versuchte, heute elementare Stoffwechselprozesse darzustellen. Als Beispiel möchte ich hier eine Elektronentransportkette, wie sie heute bei der Photosynthese und ihrer Gegenreaktion, der Atmungskette vorkommt (Abb. 1.3): Oparin ersann ein Tröpfchen, in das er Chlorophyll gab, und dem er Ascorbinsäure (Vitamin C) und oxidiertes Methylrot als Substrate anbot. Die Ascorbinsäure kann das Methylrot allein nicht reduzieren, doch wenn man das Tröpfchen mit Licht bestrahlt, gibt das Chlorophyll zwei durch Licht aktivierte Elektronen an das Methylrot ab, die Protonen folgen, und die beiden Elektronen werden der Ascorbinsäure abgezogen. So ist es am Ende doch die Ascorbinsäure, die das Methylrot reduziert. Bei der Photosynthese wird auf diese Weise NADP+ zu NADPH+H+ durch Wasser (H20) reduziert.
Sidney W. Fox: Mikrosphären
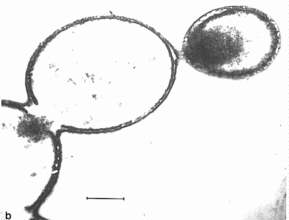
Fox kam mehr oder weniger durch Zufall zu den Koazervaten: Er experimentierte mit trockenen Aminosäuregemischen und löste sie in Wasser. Die proteinartigen Produkte haben einen Durchmesser von etwa 1-2 µm. Diese Tröpfchen entwickeln eine Doppelmembram um sich gegen die Umwelt abzugrenzen, diese ähneln zwar denen heutiger Zellen, aber unsere Membranen bestehen aus Lipid- nicht aus Proteinmolekülen. Wenn die Tröpfchen mehr Proteine aus der Lösung aufnehmen, können sie wachsen, werden sie zu groß (>15µm), teilen sie sich in einer den Bakterien ähnlichen Art und Weise.
Die Bildung von Mikrosphären erfolgt auf der Basis der physikalischen Chemie, dass hydrophobe (wasserabstoßende) Moleküle unter Wasserausschluss die maximal mögliche Anzahl an schwachen Wechselwirkungen ausbilden wollen. Daher bildet sich eine doppelte Membran. Die Zusammensetzung dieser Mikrosphärenmembran hängt entscheidend von der Ausgangskonfiguration der Aminosäuren ab, manche Aminosäuren haben eine hohe Chance eingebaut zu werden, z.B. Asparaginsäure und die nachfolgenden Aminosäuren ergeben sich aus der Startsequenz des polymers und ihren Eigenschaften als Monomere. Bei der Verknüpfung werden gleichartige Verknüpfungen (Peptidbindungen) ausgewählt, weil sie dem System einen energieärmeren Zustand ermöglichen.
Kleinere Mikrosphären können miteinander fusionieren und dadurch "mischerbig" werden, d.h. andere Strukturen ausbilden. In wässriger Lösung sind sie sehr stabil; sie werden aber instabiler und leichter verformbar, wenn sich in die Doppelmembran Lipidmoleküle einlagern.
Oparin gab in seine Koazervate Enzyme hinein, Fox' Mikrosphären wirkten von sich aus enzymatisch, da eigentlich jedes Molekül mit verschiedenartig gelagerten und geladenen Seitenketten katalytisch wirkt. So bauen diese Mikrosphären Glucose ab oder verhalten sich wie Esterasen oder Peroxidasen. Durch die Umlagerung von Seitenketten kann es hier auch zu verbesserten Enzymstrukturen kommen, sodass gewisse reaktionen zielgerichteter katalysiert werden können.
Fazit
Zusammenfassend können wir nunmehr konstatieren, daß sich aus Proteinoiden (und Proteinen) unter Mitwirkung anderer Moleküle wie Ionen, Lipiden u.a. zellähnliche Formen bilden konnten, in deren Innerem und/oder an deren Oberfläche eine Anzahl biochemischer Umsetzungen katalysiert wurden. Darüber hinaus erwarb die membranähnliche Außenschicht eine Selektivität in bezug auf Durchlässigkeit (selektive Permeabilität) bestimmter Moleküle bei Ausschluß anderer. Durch Fusion von Mikrosphären unterschiedlicher Zusammensetzung und Funktion können Fusionsprodukte entstanden sein, die die Leistungen der beiden Ausgangsformen in sich vereinigten. Gleichzeitig zeigte sich dadurch aber, daß das Auftreten, die Stabilität und die Leistungen der Mikrosphären einer Selektion unterworfen sind.
Beide Ergebnisse sind nur Analogien von Lebewesen, die gewisse Stoffwechselmodelle darstellen. Hier geht es eher darum zu zeigen, wie diese grundlegenden Funktionen auf allgemeine Vorgänge der physikalischen und organischen Chemie zurückgehen. Außerdem erkennt man hier einen chemischen Selektionsdruck, der damals wohl als einzige Form der Selektion geherrscht haben kann: Nur die Tröpfchen, die durch einen leistungsfähigen Stoffwechsel in der Lage waren zu wachsen und die Stoffe produzierten, die ihre Abgrenzung gegen die umgebende Medium verstärkten, konnten länger als die anderen überleben.
Entstandenes Leben?
Stellen die vorgestellten Strukturen ein lebendes System dar? Die Antwort darauf ist ein klares Nein, obwohl sie über eine Vielzahl von Eigenschaften verfügen, die für lebende Zellen typisch sind: Katalyse, Kooperativität (allosterisches Verhalten), Bildung von Quartärstrukturen (self-assembly), zellähnliches Aussehen, Anordnung von Molekülen zu membranähnlichen Schichten, Teilungskompetenz, Fusionskompetenz, selektive Permeabilität, Selektion. Es fehlt aber die Fähigkeit zur identischen Reduplikation, sich also fortzuentwickeln. Das wiederum beruht auf der Unfähigkeit der Proteine, die in ihnen enthaltene Information zu speichern und weiterzugeben. Man könnte sich nunmehr einen Zusammenschluß mehrerer katalytischer Aktivitäten vorstellen, der zur Autokatalyse einer jeden der daran beteiligten Komponenten führt. Die Reaktionspartner wären dann Glieder (Elemente) eines Kreisprozesses (eines Zyklus), der sich als Ganzes replizieren könnte. Das Replikationsvermögen wäre also eine Systemeigenschaft, weil keines der Glieder alleine hierzu in der Lage ist. Die Konsequenz wäre aber auch, daß das System keinerlei Änderungen (Fehler, Mutationen) zulassen würde, da durch Mutation eines seiner Elemente der darauffolgende Katalyseschritt beeinträchtigt wäre und alle nachfolgenden Schritte unterbleiben würden. Der Zyklus würde in sich zusammenbrechen (er würde aussterben). Andererseits hätte auch die Mutante praktisch keine Chance, sich zu manifestieren, denn sie müßte geeignet sein, einen völlig neuartigen Zyklus zu initiieren; die Erfolgswahrscheinlichkeit hierzu ist beliebig gering.
Zurück | Vorwärts