Ganz pauschal lässt sich sagen, dass Lebewesen nur dann miteinander arbeiten, wenn es beiden Nutzen bringt. Das ist bei den heutigen Symbiosen so. Damals brauchte man aber eine stärkere Kopplung, die RNA-Quasi-Spezies mussten nahezu vollständig voneinander abhängig sein. Diese Abhängigkeit wurde dadurch gegeben, dass eine Quasi-Spezies einer anderen die Replikation ermöglichte, ein für die andere Spezies replizierendes Enzym aktivierte oder einfach die Replikation erleichterte oder beschleunigte.
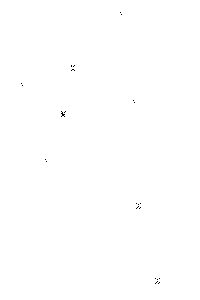
 Aus der Abbildung ist die Funktionsweise und die Kopplung innerhalb eines einfachen Hyperzyklus' ersichtlich. Das Übersetzungsprodukt der RNA I1 (E1) fördert die Replikation der RNA I2. Das Protein E2 wiederum fördert die Replikation von I1.
Aus der Abbildung ist die Funktionsweise und die Kopplung innerhalb eines einfachen Hyperzyklus' ersichtlich. Das Übersetzungsprodukt der RNA I1 (E1) fördert die Replikation der RNA I2. Das Protein E2 wiederum fördert die Replikation von I1.
Solch ein Hyperzyklus ist eine Autokatalyse zweiter Ordnung, sozusagen eine doppelte Rückkopplungsschleife. Hier sieht man, dass keine der beiden Quasi-Spezies (I1 und I2) ohne die andere existieren kann. Es kommt allerdings auf die Geschwindigkeit der Replikationen an, in welcher Konzentration die beiden Partner vorliegen, doch zerstören kann diesen Hyperzyklus nur eine Fluktuationskatastrophe, d.h. ein oder beide Partner werden räumlich voneinander getrennt, oder eine grundlegende Änderung der Umweltbedingungen, d.h. dass z.B. die Temperatur zunimmt und entweder die RNAs selbst zerfallen oder die Translationsprodukte unwirksam werden.
Hyperzyklische Kopplungen haben in Experimenten gezeigt, dass sie es schaffen, die Fehlerschwelle zu überspringen und so eine Weiterentwicklung der Quasi-Spezies möglich machen. Hyperzyklen dienen also auch dazu, dass die Gene (RNA) ein replikationsförderndes Urteil über ihre Produkte erhalten.
Ein Beispiel: Wenn, und nur wenn, unsere RNA I1 ein gutes Enzym produziert, dann vermehrt sich RNA I2 und produziert ihrerseits ein Enzym, dass I1 besser vermehrt. Würde das Enzym E1 also I2 nicht oder fehlerhaft vermehren, bleibt das replikationsfördernde Enzym E2 aus und I1 würde aussterben.
Hyperzyklen haben aber auch einen Effekt auf die Selektion. Die ersten Hyperzyklen waren wahrscheinlich noch schwach (kaum "bahnbrechende" Enzyme) und verwickelt (viele Quasi-Spezies beteiligt). Die Evolution musste diese Hyperzyklen weiter ausformen und effektiver machen.
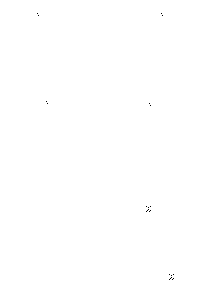
Während innerhalb von Quasi-Spezies und zwischen nicht-gekoppelten Quasi-Spezies drei Kriterien für die Durchsetzungsfähigkeit verantwortlich sind (Geschwindigkeit und Genauigkeit der Replikation, sowie Stabilität), werden in einem Hyperzyklus andere Ansprüche an die Moleküle gestellt: die RNA, die es am besten schafft, von dem sie replizierenden Enzym möglichst schnell und genau kopiert zu werden, ist die tauglichste. Durch die Bildung von RNA-Mutanten in einem Zyklus können außerdem neue katalytische Verbindungen (Strukturen und Enzyme) ausprobiert werden, was den Hyperzyklus auf eine höhere Entwicklungsstufe stellen, ihn also leistungsfähiger machen kann (s. Abb. 2).
Höherentwicklung bei Hyperzyklen
Bei Hyperzyklen treten mehrere Möglichkeiten der Weiterentwicklung ein. In der linken Abbildung kann man die allgemeine Konstellation erkennen, wenn eine Mutante schneller oder gleich schnell repliziert wird. Ist eine Mutante entstanden, die langsamer als die Stammsequenz vermehrt wird, verändert das den Hyperzyklus nicht, da beide Sequenzen dann im Zuge ihrer Selbstreplikation miteinander konkurrieren und nur die Stammsequenz schnell genug repliziert wird um den Kampf zu gewinnen.
In den rechten beiden Abbildungen wird der Unterschied zwischen phäno´- und genotypischen Veränderungen einer RNA veranschaulicht und die weitere Entwicklung des Hyperzyklus beschrieben.
 |
 |
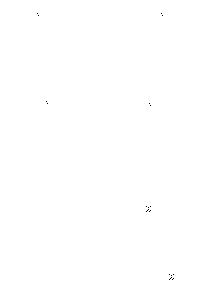 |
| Hier ist eine Mutante I1' entstanden, die von E2 besser repliziert wird als I1. Ab hier gibt es zwei Möglichkeiten: | I1' hat einen phänotypischen Effekt, wenn sie eine günstigere Zielstruktur hat und damit schneller von E2 repliziert wird. | Die Mutante I1' hat einen genotypischen Effekt, wenn sie ein Enzym (E1') hervorbringt, das I2 schneller (fehlerfreier) repliziert als E1. Hier kann allerdings I1 nicht ausgesondert werden, weil es gleich schnell repliziert wird. Eine Auswertung der Gene einer RNA ist im Hyperzyklus also nicht möglich. |
| a) E1' ist für I2 nützlicher als E1, dann wird I1 verdrängt b) E1' ist für I1 nützlicher als für I2, dann wird der Hyperzyklus auf drei Partner erweitert. c) E1' vermehrt I2 nicht mehr oder fehlerhaft, dann stirbt der Hyperzyklus aus |
||
Der Übergang aus einer oder mehreren Quasi-Spezies zum Hyperzyklus vollzog sich sicherlich ganz allmählich. Sehr primitive Translationsmechanismen, die entstanden sein mussten und die wahrscheinlich nur zum Triplett eine Aminosäure zuordneten, produzierten irgendwann leistungsfähigere Proteine als in der Ursuppe vorhanden waren. Anfangs waren diese Proteine so unspezialisiert, dass sie vermutlich ziemlich alle RNAs replizierten. Aber irgendwann drifteten die Enzymstrukturen und RNA-Strukturen so auseinander, dass es höchst wahrscheinlich war, dass zwei verschiedene Sequenzen (Quasi-Spezies) sich eher "überkreuz" Hilfe gewährten als direkt mit ihrem Produkt rückzukoppeln. Dadurch aber konnten sich die Enzyme weiter von ihren ursprünglichen Strukturen entfernen und die Kompatibilität zwischen ihnen und einer RNA schränkte sich immer weiter ein. Bis heute ist das Schlüssel-Schloss-Prinzip für Enzyme und Substrate erhalten geblieben, d.h. ein Enzym hat meist GENAU ein Substrat.
Durch diese Entwicklung nahmen auch die Hyperzyklen neuere Formen an, da immer mehr Partner aufgrund von Inkompatibilität herausfielen und andere hinzugezogen wurden.
Wie man in der Abbildung 3.2
sehen kann, haben Quasi-Spezies und die resultierenden Hyperzyklen einen großen Nachteil: Ausschließlich die phänotypischen, d.h. strukturellen Eigenschaften einer RNA (Replikationsmerkmale und Stabilität) werden durch die Selektion bewertet. Die Gene selbst, die ja die Enzyme kodieren, können nicht selektionell erfasst werden, die Enzyme verbessern sich dadurch nicht stetig, sondern zufällig durch Mutationen.Wie dieses Problem gelöst wurde, lesen Sie im nächsten Kapitel.
Zurück | Vorwärts